












【佳学基因检测】并指症基因检测应该怎么做?如何避免假阴性结果?
并指基因检测的位点
并指畸形(syndactyly)是一种手足畸形,其特征是两个或多个手指或脚趾未完全分开。此病症可分为综合征性和非综合征性两大类,其中涉及的基因和位点众多。科学研究已识别出多个与并指畸形相关的关键基因,这些基因位点对于诊断和筛查具有重要意义。
-
GJA1基因:该基因编码连接蛋白(connexin 43),主要参与细胞间的电信号传递。GJA1突变与非综合征性并指畸形相关,并可能影响手指和脚趾的分化过程。
-
HOXD13基因:HOXD13基因在胚胎发育中扮演重要角色,尤其在四指和脚趾的发育过程中。HOXD13突变与非综合征性并指畸形有关,它可能通过调节皮肤和骨质发育的途径来影响并指畸形的发生。
-
FGF16基因:纤维母细胞生长因子16(FGF16)与胚胎发育中的组织模式形成有关,它的突变可能导致手指或脚趾的分化异常。
-
LMBR1基因:LMBR1基因与综合征性和非综合征性并指畸形都有关。该基因在胚胎发育中调控四指网的形成,其突变可能影响四指的正常发育。
-
LRP4基因:LRP4基因在与WNT信号传导相关的发育通路中发挥作用,其突变与Cenani-Lenz综合征相关,也可能影响并指的发生。
-
HOX13基因:HOX13基因也是与并指畸形相关的重要基因,尤其在综合征形式的并指中扮演重要角色。
这些基因及其位点的识别有助于提供精准的诊断和筛查方法,尤其是在产前诊断中,当传统的放射诊断方法难以实施时,基因检测提供了一个有效的替代方案。
并指基因检测的科学性
并指基因检测的科学性体现在几个方面:
-
遗传异质性:并指畸形的遗传基础非常复杂,不同基因的突变可以导致类似的表型。例如,LRP4基因突变与Cenani-Lenz综合征相关,但其他基因如APC也能导致类似的表型。这表明并指畸形的遗传机制可能涉及多个基因和信号通路。
-
非编码区域和内含子变异:基因检测不仅关注外显子中的突变,还需要考虑内含子和非编码区域中的变异,这些区域可能也会对基因功能产生影响。例如,表观遗传学的变异和非编码RNA可能会影响基因表达,进而影响并指的发生。
-
多路径通路的参与:并指畸形的发生不仅涉及WNT、Notch和ADAMTS信号通路,还可能涉及其他尚未明确的通路。因此,基因检测不仅要关注已知的关键基因,还应考虑这些基因之间的相互作用。
-
表型异质性:并指畸形的表型异质性意味着同一基因的突变可能在不同个体中表现出不同的症状或严重程度。这要求基因检测不仅要识别出突变,还需要结合具体的临床表现来解读结果。
并指基因检测的意义
并指基因检测具有重要的临床和研究意义:
-
早期诊断和干预:通过基因检测可以在产前或出生后早期诊断并指畸形,从而采取早期干预措施。这对于减少并指畸形对患者生活质量的影响具有重要意义。
-
个体化治疗:基因检测提供的详细遗传信息有助于制定个体化的治疗方案。例如,对于不同基因突变导致的并指畸形,可以根据具体的突变类型选择最适合的干预措施。
-
筛查和预防:在产前筛查中应用基因检测,可以识别出高风险的胎儿,从而提供针对性的监测和干预。特别是在高风险家庭中,基因检测可以帮助识别潜在的遗传风险。
-
研究和新治疗的开发:基因检测不仅帮助理解并指畸形的遗传机制,还有助于开发新的治疗方法和生物标志物。例如,通过研究涉及并指畸形的基因和通路,可以发现新的治疗靶点。
-
综合征的分类:在一些综合征中,并指畸形可能是分类的重要依据。例如,在Smith-Lemli-Opitz综合征(SLOS)中,第二和第三脚趾的并指可以作为诊断的关键特征。
总的来说,并指基因检测的位点和科学性对于理解和管理并指畸形具有重要的临床和研究价值。通过进一步的研究和技术进步,我们可以期待基因检测在并指畸形的早期诊断、个体化治疗和疾病预防方面发挥更大的作用。
综合征性并指遗传决定因素
根据《人体手指异常的基因原因》,发生在人身上的并指畸形有 300 多种综合征形式,因此其分类复杂,需要借助基因检测进行明确。并指症基因检测的大数据分析收录了按照发病机制进行分类的部分并指综合征的临床表征,将其列于表1。
| 基因 | 综合征类型 | OMIM | 常见变异 | 遗传方式 | 临床表现 | 如何到致疾病的发生 |
| 1 (a) 编码区域中的遗传决定因素 | ||||||
| ROR2 | 短指症 B 型(BDB1) | 113000 | c.1324C.T; p.R441X | AD | 短指症的特征是远端指骨发育不全/发育不全,伴有远端指骨联合畸形、腕骨/跗骨融合和部分皮肤并指畸形 | 促进 WNT 过度表达 |
| SOST | 硬化性骨化病 | 269500 | 无义突变 | AR | 很多病例存在食指和中指皮肤不对称并指畸形。下颌呈异常方形 | |
| LRP4 | 硬化性骨化病,2 | 614305 | c.3508C > T; p.R1170W and c.3557G > C; p.W1186S | AD, AR | 渐进性骨骼过度生长。并指畸形是一种可变的表现 | |
| GLI3 | Pallister–Hall 综合征 | 146510 | 单倍体不足,c.1468_1469insG 和 c.1007_1008dupAC | AD | 下丘脑错构瘤、垂体功能障碍、中枢性多指畸形和不同程度的并指畸形 | BMP 抑制 |
| 格雷格头颅多指并指综合征 | 175700 | c.2374C > T; p.Arg792* | AD | 与轴前和轴后多指畸形和可变并指畸形相关的额叶隆起、舟状头畸形和眼距过宽 | ||
| LMBR1 | 三指节拇指-多指并指综合征 | 174500 | ZRS 增强子上的 287 位 | AD | 这种畸形的拇指通常是对生的,并具有正常的掌骨。并指畸形的程度各不相同 | |
| DHCR7 | Smith-Lemli-Opitz 综合征 | 270400 | c.453G > A; p.W151X | AR | 影响多个身体系统,其中 2 和 3 趾并指畸形是常见症状 | |
| RAB23 | 卡彭特综合征 | 201000 | 纯合无义/移码致病变异 c.434 T > A; p.L145X | AR | 颅缝早闭、多指并指、肥胖和心脏缺陷 | |
| FGFR2 | Apert 综合征 | 101200 | c.755C > G; p.S252W or c.758C > G; p.P253W | AD | 颅缝早闭、中面部发育不全、手足并指畸形,且有骨结构融合的趋势 | FGF过度表达 |
| FGFR1/FGFR2 | 菲弗综合征 | 101600 | FGFR1 – p.P252R | AD | 1 型为经典综合征,与生活相容,包括颅缝早闭、中面部缺陷、拇指宽、大脚趾宽、短指畸形和可变性并指畸形 | |
| TWIST1;FGFR3 | Saethre–Chotzen 综合征 | 101400 | FGFR3 p.P250R, deletion of TWIST1 | AD | 颅缝早闭、面部畸形、手足畸形。并指畸形的程度也各不相同 | |
| HOXD13 | 短指并指综合征 | 610713 | 多聚丙氨酸收缩,c.950A > G;p.Q317K | n.r | 短指畸形和并指畸形(部分皮肤织带)与少指畸形相关 | 视黄酸抑制 |
| LRP4 | 硬化性骨化病2型 | 614305 | c.1151A > G; p.Tyr384Cys | AD, AR | 渐进性骨骼过度生长。并指畸形是一种可变的表现 | Notch 信号的抑制 |
|
其他基因位点/基因突变序列 |
临床表现 | 如何到致疾病的发生 |
| 1 (b) 非编码区域中的遗传决定因素 | ||
| 内含子 EMID2 | 全前脑畸形谱系障碍和严重上肢并指畸形 | 异位SHH表达 |
| 内含子 5 LMBR1 | 无多指畸形的皮肤并指畸形 | ICD 减少 |
| 内含子IRF6 | 范德沃德综合征(MIM:119,300) | 未分类 |
| 22 个内含子YY1AP1 的外显子化 | Grange 综合征(MIM:602,531)伴有有效性皮肤并指畸形。手指:右手第三、第四、第五根手指和第三/第四根手指。脚趾:第二/第三根脚趾双侧皮肤并指畸形 | 未分类 |
| 内含子 6 KATNB1 | 先天性小头畸形、无脑畸形、身材矮小、多指畸形和牙齿异常 | 未分类 |
| 内含子 8 FGFR2 | Apert 综合征(MIM 101,200) | 未分类 |
| IVS8-1G > C 基因突变序列DHCR7 | Smith-Lemli-Opitz 综合征(MIM 270,400) | BMP 抑制 |
| 前 ZRS 区域 | TPS(MIM:174,500) | 异位表达 |
AR常染色体隐性遗传,AD常染色体显性遗传,XLR X 连锁隐性遗传,nr由于证据不足未报告,EMID2 EMI 结构域含 2,LMBR1肢体发育膜蛋白 1,IRF6干扰素调节因子 6,YY1AP1 YY1 相关蛋白 1,KATNB1 Katanin,p80 亚基 B1,FGFR2成纤维细胞生长因子受体 2,TPS拇指多指并指综合征
编码区域的变异
下面列出了一些综合征及其在典型发病机制中每个相互作用的记录,但必须注意的是,还存在许多其他综合征。每种综合征的遗传变异列于表格1。
观察到的一些主要 WNT 过度表达综合征包括 B 型短指症 (BDB1) (MIM 113000)、硬化性骨化病 (MIM 269500) 和硬化性骨化病 2 (MIM 614305)。
BDB1 被定位到ROR2基因 (9q22.31),该基因编码 WNT 配体(如 WNT5a)的辅助受体。由于已发现 ROR2 的 CRD 与 Frizzled 受体的 WNT 结合域惊人地相似,因此推测 ROR2 可能与经典的 WNT 信号转导有关。它也被认为参与了非经典的 WNT-PCP 相互作用,因为与 WNT5a 突变小鼠相比,具有突变 ROR2 的小鼠产生相似的表型。硬化性骨化病 2 已被定位到LRP4基因,其中硬化素和 LRP4 之间存在直接相互作用,并且 LRP4 促进硬化素介导的 WNT 抑制;这与LRP4突基因突变序列有关。此外,已知 LRP4 与 Notch 通路的相互作用在进化上是保守的,并且LRP4基因中高度保守的 EGF-2 钙结合结构域中的纯合错义基因突变序列也被推测可以消除 Notch 信号传导。
贼后,在骨硬化症中,功能丧失的SOST基因变异可能使硬化素抑制 LRP5/6 和卷曲受体复合物,从而导致 β-catenin 磷酸化和下调,从而抑制 WNT 信号相互作用。
被认为促进 BMP 抑制的主要综合征包括 Pallister-Hall 综合征 (PHS) (MIM 146510) 和 Greig 头颅多指并指综合征 (MIM 175700)、三指节拇指多指并指综合征 (TPS) (MIM 174500)、Smith-Lemli-Opitz 综合征 (SLOS) (MIM 270400) 和 Carpenter 综合征 (MIM 201000)。PHS 和 GCPS 均与GLI3基因相关。然而,Pallister-Hall 综合征与中枢性多指有关,而 GCPS 涉及前轴和后轴性多指。GLI3基因(7p14) 编码一种转录因子,该转录因子是 SHH 信号的双功能下游调节剂;与这两种综合征都有关。大多数GLI3基因变异与GLI3基因单倍体不足有关,贼终导致 GLI3 激活剂 (GLIA) 或阻遏物 (GLIR) 形成的偏差。TPS 的常见遗传决定因素包括跨越LMBR1基因的重复和RAB23基因(6p12.1-q12),该基因编码音猬因子 (SHH )信号的负调节剂,在 Carpenters 综合征中受到干扰。对于 SLOS,已知编码酶 7-脱氢胆固醇 (7-DHC) 还原酶 ( DHCR7 : 11q13.4) 的基因被失调。据推测, DHCR7基因协调胆固醇部分和 SHH 信号传导,从而控制并指表型的严重程度。
已证实可促进 FGF8 过度表达的综合征性指症主要包括 Apert 综合征 (MIM 101200)、Pfeiffer 综合征 (MIM 101600) 和 Saethre-Chotzen 综合征 (MIM 101400)。FGFR2 基因变异表明该受体在 Apert 综合征和 Pfeiffer 综合征中均发挥作用。P253W 突基因突变序列可能会扰乱对 FGF10 的配体结合特异性,导致外胚层中的 FGF10-FGF8 环过度刺激 FGF8。对于 Saethre-Chotzen 综合征,TWIST1和FGFR1/3基因已被定位到该综合征。TWIST1被归类为 FGF 的上游调节剂,因此调节 FGF 表达。
贼后,短指-并指综合征 (MIM 610713) 与HOXD13基因变异 (多聚丙氨酸收缩和错义变异) 有关。据推测,HOXD13突基因突变序列会抑制ECM 中的视黄酸 (RA)。
非编码区域的变异
筛查该基因及其侧翼区域的重要性仍然至关重要,因为非编码区域中的替代剪接基因突变序列可能导致外显子跳跃和表观遗传影响,从而导致蛋白质畸形。修饰基因的存在被认为会影响p63 和RAB23 基因基因突变序列的表型,这可能是造成家族内变异的原因。许多内含子位点(表(表格1),1目前,已有 150 多个异常信号通路在并指表型中被发现,但大多数异常信号通路需要对其进行分类。
非综合征性并指遗传决定因素
根据表型多样性,非综合征性并指畸形可分为 9 种类型(I-IX)(表(表2),2), 以及相应的基因/位点。已发现的一些基因包括HOXD13、FBLN1、GJA1、LMBR1、LRP4、GREM1(BMP 拮抗剂)、FGF16以及贼近发现的BHLHA9。
表 2:与非综合征性并指畸形相关的假定发病机制的基因/位点
|
非综合征性并指畸形的类型 |
基因座/基因 | OMIM | 常见基因突变序列 | 遗传方式 | 临床表现 | 发病原因 | |
| I-a | ZD1;对指畸形;Weidenreich 型 | 3p21.31 | 609815 | n.r | AD | 双侧、对称、 | n.r |
| 手指:正常 | |||||||
| 脚趾:仅 2/3 | |||||||
| I-b | SD1; Lueken型 | 2q34-q36 | 185900 | n.r | AD | 通常,双侧 | n.r |
| 手指:3/4 手指,皮肤/骨质 | |||||||
| 脚趾:2/3 脚趾,皮肤 | |||||||
| I-c | 蒙塔古型 | 2q31-q32 | n.r | c.917G > A; p.R306Q, | AD | 通常情况下,双边 | 抑制视黄酸 |
| HOXD13 | c.916C > G; p.R306G | 手指:仅限 3/4 手指,皮肤/骨骼 | |||||
| 脚趾:正常 | |||||||
| I-d | 卡斯蒂利亚型 | n.r | n.r | n.r | AD | 双边 | n.r |
| 手指:正常 | |||||||
| 脚趾:4/5 仅脚趾,皮肤 | |||||||
| II-a | SPD1;沃尔丁堡型 | 2q31; | 186000 | Polyalanine repeat expansions, frameshift deletions, 2q31.1 microdeletion and G11A missense | AD | 手指:SPD,中轴(3/4 手指) | 抑制视黄酸 |
| HOXD13 | 脚趾:SPD,轴后(4/5 个脚趾) | ||||||
| II-b | SPD2;德比尔型 | 22q13.3; | 608180 | t(12;22) (p11.2;q13.3) | AD | 手指:SPD 位于中心和轴后 | FGFR8 表达增加 |
| FBLN1 | 脚趾:轴后并指畸形 | ||||||
| II-c | SPD3; Malik type | 14q11.2-q13 | 610,234 | n.r | AD | 手指:SPD是核心 | n.r |
| 脚趾:SPD 后轴 | |||||||
| III | SDTY3;Johnston-Kirby型 | 6q21-q23; | 186100 | nt427G > A and | AD | 手指:4/5 根手指有效双侧并指;第五根手指短小,中指缺失或发育不全 | 降低下游 BMP2 表达,导致 FGF4 和 FGF8 过度表达 |
| GJA1 | c.T274C; p.Y92H | 脚趾:正常 | |||||
| IV-a | SDTY4;哈斯型 | 7q36; ZRS (LMBR1) | 186200 | SHH 调控元件 (ZRS) 中的杂合变异和重复 | AD | 有效性、双侧性,常伴有多指 | 改变 ZRS 控制/肢体特定的 SHH 表达 |
| 手指:所有手指均有蹼;前轴/后轴多指,杯状手 | |||||||
| 脚趾:正常 | |||||||
| IV-b | 安德森-汉森型 | n.r | n.r | n.r | n.r | 手指:所有手指均有蹼;前轴/后轴多指,杯状手 | n.r |
| 脚趾:趾蹼多变,伴有多指畸形 | |||||||
| V | SDTY5;Dowd型 | 2q31; | 186300 | c.950A > G;p.Q317R,以及多聚丙氨酸扩增 | AD | 有效的 | 抑制视黄酸 |
| HOXD13 | 手指:4/5 手指掌骨融合;掌骨发育不全 4/5 | ||||||
| 脚趾:中轴蹼 | |||||||
| VI | 连指手套类型 | n.r | n.r | n.r | AD | 单侧的 | n.r |
| 手指:2/5 手指 | |||||||
| 脚趾: 2/5 脚趾 | |||||||
| VII-a | Cenani-Lenz 型;勺手型 | 11p12–p11.2; | 212780 | 错义变异贼为常见,c.1117C > T;p.R373W | AR | 手指:全骨性并指,掌骨融合,匙头形状 | WNT 过表达 |
| LRP4 | 脚趾:全骨性并指畸形,伴有跖骨融合 | /推测会导致 Notch 信号抑制 | |||||
| VII-b | 少指畸形 | 15q13.3; | n.r | GREM1和FMN1 1 基因存在 1.7 Mb 重复 | AD | 手指:少数手指变形 | BMP 拮抗剂 GREM1(通常受 FGF 抑制);在突变形式下活跃,抑制 BMP 活性 |
| GREM1-FMN1 | 脚趾:趾骨多变 | ||||||
| VIII-a | Orel-Holmes type | Xq21.1; FGF16 | 309630 | 外显子 3 中的无义基因突变序列 p.R179X 和 p.S157X | X-R | 手指:4/5 掌骨融合 | 损害 FGF16-FGFR1 相互作用 |
| 脚趾:正常 | |||||||
| VIII-b | 勒奇型 | n.r | n.r | n.r | AD | 手指:4/5 掌骨融合 | n.r |
| 脚趾:正常 | |||||||
| IX | MSSD;Malik-Percin 类型 | 17p13.3/ | 609432 | c.311T > C; p.Ile104Thr | AR | 手指:中轴骨性并指畸形,伴有指骨退化 | 异位 FGF8 表达限制 BMP 抑制 |
| BHLHA9 | 脚趾:前轴蹼;远端指骨发育不全 | ||||||
然而,关于非综合征性并指症分类也存在一些争议。多指并指基因解码的收录数据中有一例皮肤并指症,发生在二/五指或三/五指中,而不伴有多指畸形。这种不伴有多指畸形的孤立性并指症类似于 III 型并指症,后者是由GJA1基因变异引起的,已在四/五指或三/五指中观察到。然而,尽管与 III 型相似,但作者观察到 ch7q36.3( LMBR1区域)中存在一种变异,该变异与具有相似表型的小鼠的锤状趾基因座同源。这可能突显出这是一种遗传异质性的病例,并支持这种类型可能是多基因的观点。
另外两份关于 IX 型并指畸形的报告以第三和第四掌骨骨性愈合为特征,由此相关的表型略有不同。佳学基因致病基因鉴定基因解码收录了一例涉及第二和第三近端指骨的 MSSD 病例,这与 Malik 的原始分类不同,并将其归因于一种新的无综合征类型。然而,Malik不同意这一观点,并将其重新归类为与BHLHA9基因相关的无综合征 IX 型。另有一个基因检测案例属于MSSD 病例,其第二和第三掌骨近端指骨呈三角形,与 Weinrich 的病例报告相似,此外还有尺骨半肢畸形(缩短),这是一个新发现 。这两例 MSSD 病例均未进行基因检测,分类结论基于手部 X 光检查。如果它映射到同一个基因,这可能是与BHLHA9基因相关的一种表型异质性。因此,基因检测仍然是必不可少的,以便能够进一步评估这些不一致之处。
应用于基因筛查
有关并指畸形基因的新证据可能提供一种替代的基因筛查方法,尤其是在几乎无法进行放射诊断的产前诊断中。非综合征性并指畸形所涉及的GJA1、HOXD13和FGF16基因涉及第四指网,可以以产前筛查为借口提供诊断选择,以防止以后出现成角畸形 。LMBR1 、LRP4和HOX13基因在综合征和非综合征形式中均被失调(图 1)。此外,HOXD13可能能够调节皮肤和骨质并指通路。因此,将这些基因纳入基因组将扩大诊断标准。在诊断过程中,如果基因检测未得出结论,则筛查侧翼区域和内含子区域可能很重要。不要忘记,在综合征中,并指本身的存在有时可能是对综合征进行分类的决定性因素(例如:SLOS 中的第二/第三脚趾并指)。
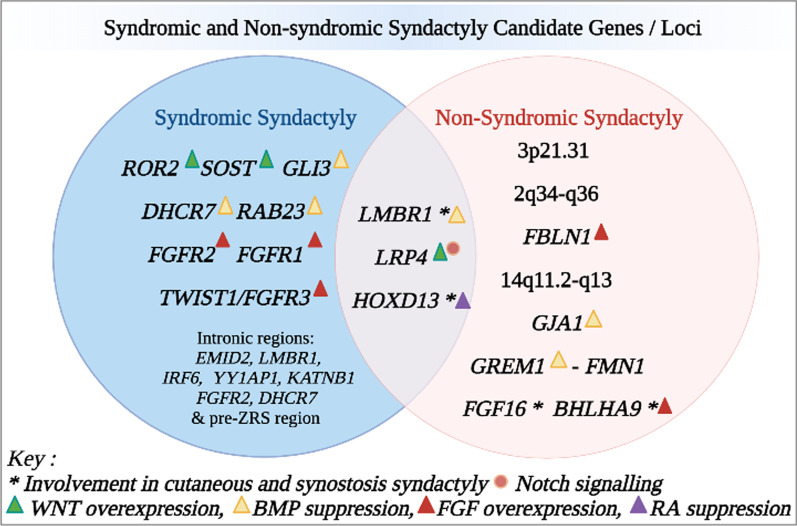
图1:多指并指基因解码基因检测分析了综合征性和非综合征性并指症的基因和基因位点的代表性。这些变异根据其参与记录的并指症发病机制相互作用而进一步表征。WNT无翅型整合位点家族、BMP骨形态发生蛋白、FGF 成纤维细胞生长因子、RA视黄酸
进一步明确并指发生的基因原因
非综合征性并指症被认为具有很好的特征,但相关的 MSSD 表型存在争议,两例病例报告均未包括 BHLHA9 的基因检测,因此不能排除表型异质性的可能性。遗传异质性;尽管 Cenani Lenz 表型被归因于基因LRP4,但也有基因解码称APC基因具有相同的表型。此外,Qattan 的研究中 III 型并指症可能证明除了GLI3之外还有另一个基因座。此外,由于 BMP 是分泌因子,基因表达模式不会与功能观点有效相关。因此,还需要考虑蛋白质分布。此外,了解基因-基因相互作用、非编码变异和表观遗传学有助于更好地表征并指表型。
对于第一和第四蹼状并指畸形,建议在婴儿 6 月龄左右时尽早分开。因此,除了涉及这些蹼状并指畸形的其他已知基因外,可以优先检测与无综合征 Castilla 型和 Lerch 型相关的基因。并指畸形的复杂性进一步增强,因为一个基因可能涉及多条通路。已知LRP4基因基因突变序列与 WNT 的过度表达有关,但也有基因突变序列显示 Notch 信号传导被破坏。另一方面,大多数综合征并指表型在可能参与的通路方面并没有明确下来,这是因为很多基因检测没有采用更为先进的基因解码技术。
孟德尔性状和全基因组关联研究(GWAS)的治疗靶点更有可能在多因素疾病的临床研究中取得成功。因此,在这方面进行基因检测分析对于医生和患者来说都有明确的治疗价值。
并指基因检测的共识性意见
手足的自然发育是一个受到严格调控的过程,涉及大量基因。传统观点认为,WNT 经典通路导致趾间细胞死亡,从而导致并指畸形。然而,许多信号传导通路还未明确的基因也参与了这一疾病过程的发生,大多数其他通路仅在动物研究中进行研究。并指发生的致病基因鉴定基因解码认为,经典和非经典 WNT 信号、Notch 信号、ADAMTS 金属蛋白酶通路和非编码区域中的基因聚集可能协同作用,以建立正常的肢芽发育。对HOXD13、LMBR1、FGF16、BHLHA9进行候选基因研究,以了解它们对皮肤和骨性并指畸形表型的贡献,可以增强筛查。由于迫切需要进行产前基因检测,因此GJA1、HOXD13和FGF16基因以及涉及第一和第四个蹼的其他基因也可能被优先考虑。总的来说,将这些发现转化为人类研究,尤其是通路图谱,可以进一步解决与并指畸形相关的遗传基础。此外,它将有助于治疗并指畸形并促进生物标志物的发现。
(如果您已经做了基因检测,想获取与基因检测型相对应的治疗方案,请点击此处上传您的基因检测结果)
(责任编辑:佳学基因)
